| Источник: SciTecLibrary.com Авторы: Прангишвили И.В., акад. АН Грузии, докт. техн. наук, Гаряев П.П., акад. РАМТН, докт. биол. наук, Тертышный Г.Г., канд. техн. наук, Леонова Е.А., Мологин А.В.
Генетические структуры как источник и приемник голографической информации. Институт проблем управления РАН, Москва.
Живое и Неживое имеют общие характеристики информационно-голографического характера. Это связано с фундаментальной способностью когерентных физических полей интерферировать и, тем самым, при наличии регистрирующей среды, “записывать” собственные характеристики. Существенно, что в этом процессе не играет роли какие структуры, Живые или Неживые, порождают эти поля. Развивая эти идеи, опубликованную нами ранее [1, 2], в настоящей работе мы уточняем нашу гипотетическую модель эндогенного биоволнового (голографического) управления развитием многоклеточных организмов из эмбрионального состояния во взрослое. Кроме того, в последние годы выясняется, что к числу фундаментальных явлений природы относится так называемый “фоновый принцип” [3], непосредственно связанный с биоголографической работой генетического аппарата [1, 2]. Суть фонового принципа заключается в том, что известное соотношение шум/сигнал трактуется так, что уровень сигнала можно повысить не за счет подавления шума (фона), как это делается обычно, а наоборот, фон служит источником сигнала и средством повышения его уровня. Этот принцип универсален и работает во всех областях человеческого знания, в общем отображая категории Части и Целого, в том плане, что полное знание о части невозможно без учета свойств целого. Например, в лингвистике фоновый принцип хорошо виден в случае омонимий, когда слова-омонимы (‘лук’ ‘рейд’ ‘коса’ и т.д.) не могут иметь точную семантику вне целого предложения, то есть контекста (фона). В генетической лингвистике применение фонового принципа позволило нам решить [2] парадокс Вобл-гипотезы Ф.Крика, в соответствии с которой в генетическом коде существует задача снятия омонимических неоднозначностей знаковых дублетов кодонов. Эту проблему живая клетка, точнее ее белок-синтезирующий аппарат, снимает именно на основе фонового принципа за счет контекстных ориентаций рибосомы в геометрически одномерном смысловом пространстве информационных РНК. В свою очередь, такие контекстно-фоновые механизмы “логической” ассоциативной работы рибосомы невозможны без интегрально-волнового восприятия целого генетического текста информационных и, возможно, преинформационных РНК. Другой вектор приложения логики фонового принципа заключается в том, что в радиоволновом диапазоне можно обнаруживать и опознавать макрообъекты-“невидимки”, изготовленные по технологии “Стелс”. Фоновый принцип делает вполне реальной разрабатку микроскопии (в дополнении к фазово-контрасной) прозрачных для света живых и динамичных биоструктур без повреждающей предварительной окраски. Можно наблюдать, например, сложную знаковую динамику хромосом в живых клетках и связать ее с волновыми управляющими функциями генетического аппарата, а это до сих пор было недоступно для исследователей. Известно, что хромосомный континуум многоклеточных организмов является высокоорганизованной системой генерации эндогенных когерентных электромагнитных полей широкого спектра от 250 нм до 800 нм, далее ИК- и СВЧ-диапазоны [1, 2]. Высшим биосистемам свойственна также знаковая акустика генетических структур, которая может иметь солитонный и когерентный характер. Эти информационно-волновые процессы распространяются и на белковую, и на водную составляющие биосистем [1, 2]. Поэтому практически любая точка в пространстве организмов является источником физических полей, в том числе генознаковых. Все организмы земли взаимодействуют с внешними естественными и искусственными электромагнитными и акустическими полями, некоторые из которых могут быть когерентными, например, поля от таких удаленных источников, как звезды. Имея в виду эти соображения, применим принцип Гюйгенса к биосистемам и будем считать, что каждая точка живой клетки является источником элементарной электромагнитной и/или акустической волны. Комплексная амплитуда волны в каждой такой точке равна сумме комплексных амплитуд отдельных элементарных волн, излучаемых каждой точкой биосистемы (далее речь идет только об электромагнитных волнах). Поскольку нас, прежде всего, интересуют генетические волновые процессы, будем рассматривать в качестве источников элементарных волн управления жидкокристаллические фазы ДНК в составе хромосомного континуума. Характерным для таких фаз как in vitro так и in vivo является их сложная высокоорганизованная надмолекулярная слоистая структура, состоящая из холестерических сверхплотных укладкок спиралей ДНК [4]. Такие слои ДНК можно рассматривать как аналоги транспарантов в голографии. Физико-математическая модель процесса излучения волны молекулами ДНК записывается следующим образом. Допустим, что в некоторой плоскости координат x,y размещен ограниченный элементарный плоский слой жидкокристаллической ДНК, слой, который образовал излучательный континуум. Тогда элементарный фронт волны, исходящей из произвольной точки x на слое ДНК, представим в виде
Суммарная
амплитуда элементарных волн в точке (1)
При интегрировании в бесконечных пределах соотношение (1) с точностью до постоянного члена перед интегралом является формулой дифракции. Ввиду того, что расстояние rs есть иррациональная функция разности координат, ограничимся небольшой областью в окрестности оси z и применим параболическое приближение (2)
rs = Zs + Отличие голограммы от квазиголограммы состоит в способе их формирования (записи). Для регистрации обоих типов голограмм требуются опорные (референтные) волны. Однако, для квазиголограмм референтные волны могут быть фрагментами объектных волн и выступать как квазисферические за счет дифракции на отдельных микроструктурах биосистемы. По этой причине формируются многоопорные референтные пучки для формирования градиентов поля квазиголограммы. Такого рода квазиголограммы будем называть волновыми (в-квазиголограммы). Квазиголограммы ДНК могут формироваться и другим путем, который зависит от последовательностей нуклеотидов, определяющих характер жидкокристаллических упаковок хромосом и, следовательно, структуру (информацию) квазиголограмм [1]. Их мы будем называть последовательность-зависимыми (п-квазиголограммами). Однако, в чистом виде ‘в’- и ‘п’- типы вряд ли существуют. Скорее всего, формируются смешанные типы. Есть основания думать, что в биосистемах образуются и квазиголограммы, и голограммы [1, 2]. Поэтому, далее в статье употребление этих терминов по биологическому смыслу (конечному биорезультату) семантически тождественно. Ключевую роль в волновом управлении формирования таких голографических решеток могут играть факторы изменения градиентов напряженности физических полей, пронизывающих гетерогенное пространство организмов и квантованно (по углу регистрации-считывания, по длине волны и по субстратам записи) записывающих - воспроизводящих его динамичную структуру. Эти градиенты зависят от конкретных биоструктур и от голограмм - квазиголограмм, образуемых ими. Именно на них дифрагируют фотонные и другие поля с образованием знаково-непрерывного континуума градиентов этих полей. Градиенты в данном случае выступают как носители биознаков пространственной разметки биосистем, а совокупность градиентов образует знаковый континуум построения многоклеточного организма. Важным моментом волновой организации биосистем является фактор безынерционной радиосвязи по поляризационному каналу между ядрами клеток биосистемы для их синхронизации, введения в когерентное состояние “знания” друг о друге. Такие радиоволновые излучения могут индуцироваться лазерными излучениями хромосом по типу обнаруженного явления конверсии фотонов в радиоволны [2, 6, 7]. В тесной связи с этим ключевой (в биологическом смысле) является предполагаемая нами фрактально масштабированная когерентность (связь) по каналу поляризации фотонов с радиоволнами (Ф® Р гипотеза) на уровнях хромосомного континуума и других управляющих организмом биоструктур, таких как ферментные ансамбли, цитоскелет, внеклеточный матрикс и т.д. В рамках Ф® Р гипотезы снимается проблема радиоволновой “прозрачности” организмов и, соответственно, биологической активности радиоволн, знаково модулированных по поляризации. Действительно, существуют противоречивые данные, связанные с “прозрачностью”, т.е. отсутствием выраженного действия хаотически поляризованных радиоволн кило- и мегагерцового диапазонов на живые организмы. Можно полагать, что эти диапазоны будут так или иначе воздействовать на биосистемы, если они имеют знаконесущие поляризационные модуляции. Это означает, что они будут вызывть биологический (метаболический) ответ при наличии поляризационных резонансов с радиоизлучающими и радиопринимающими командными биоструктурами, например, с ДНК, РНК и белками. Подставив выражение (2) в (1), мы получим формулу дифракции Френеля. Дифрагирующая объектная волна “записывает” информацию о геометрии (форме) биоучастка, на котором происходит акт дифракции. В простейшем варианте это можно представить в виде ряда событий. Волна, излученная слоем хромосомной ДНК, выходя за пределы клеточного ядра, дифрагирует на встречающихся клеточно-тканевых структурах, то есть модулируется по амплитуде, фазе, поляризации, частоте и тем самым приобретает информацию о строении этих структур. Это фактор автоконтроля биосистемы за своей пространственной структурой [1, 2]. Для регистрации в форме голограммы полученной информации нужна референтная волна, которую представим как сферическую и запишем ее также в параболическом приближении (3)
Здесь (4)
В результате сложения комплексных амплитуд объектной и референтной волн получим выражения для интенсивности, равной квадрату абсолютной величины комплексной амплитуды результирующего поля (5)
Раскрывая квадрат от модуля комплексной амплитуды суммарного поля, получим (6)
Можно полагать, что, следуя закону распределения интенсивности поля или его градиенту, биосистема будет использовать эти градиенты как некоторые (из многих) векторы построения своей клеточно-голографической структуры именно в тех местах, которые соответствуют амплитудному пропусканию голограммы в соответствии с условным перераспределением интенсивности и связанным с ним ростом и развитием биоструктур, взаимодействующих с фотонами. Возникает понятие “условного потемнения”, которое мы понимаем как интерференционный результат сложения когерентного излучения по амплитуде, фазе, поляризации и частоте. Эти изменения модуляции интенсивности поля происходят вследствие сгущений и разряжений оптической плотности при динамике клеточно-тканевых структур в ходе развития и жизнедеятельности биосистем. Законы этих сгущений и разряжений и перемещений, т.е. картины и сценарии интенсивности организменных полей, в том числе их оптическая активность и спектральный состав, являются содержанием цитодифференцировок и морфогенеза организмов в волновом представлении. Заметим, что под частотной (спектральной) модуляцией фотонов, проходящей на биоструктурах, надо понимать не только и не столько известное комбинационное рамановское преобразование частот в видимом и инфракрасном интервалах длин волн. Мы допускаем, что в биосистемах может иметь место аналог феномена, обнаруженный нами ранее [2, 6, 7]. Он, в частности, проявляется в трансформации когерентного света, в определенных условиях взаимодействующего с геноструктурами и белками, в биологически активные радиоволны кило- и мегагерцового диапазонов. При этом, как уже упоминалось, существенна корреляционная зависимость поляризации фотонов и поляризации рожденных из них радиоквантов как потенциальный биоинформационный канал. Не исключено, что этот канал является модификацией открытого недавно эффекта квантовой телепортации связанных по поляризации фотонов, предсказанного в 1935 г. как эффект квантовой нелокальности (квантовой телепортации) Эйнштейна - Подольского - Розена [9]. Возможно, этот канал обеспечивает адекватное восприятие и использование биосистемами внутренней и внешней волновой среды их обитания. Для правильного понимания предлагаемой голографической модели организации биосистем, надо учитывать, что квази - спонтанная динамика биоструктур in vitro - in vivo, по крайней мере, для таких ключевых как ДНК, рибосомы и коллаген, носит нелинейный квантованный характер и реализуется в рамках явления возврата Ферми-Паста-Улама (ФПУ), т.е. является солитонным процессом с памятью на начальные условия возбуждения колебательных мод. Указанные биоструктуры, возможно, имеют квантованный спектральный состав акустических колебаний с меняющимися, но периодически повторяющимися некоторыми модами [1]. Здесь физика колебательных процессов обеспечивает фундаментальный атрибут биологической памяти генетических молекул, а именно их способность к волновому “самопрочитыванию” генетических “текстов” и голограмм с возвратами ФПУ к уже “прочитанному” и/или “просмотренному” через высокоорганизованные и квантованные вещественно-волновые состояния, как особого рода “памяти” хромосом. Именно здесь, в вещественно-волновых измерениях, заложена стратегия волнового программирования развивития и эволюционирования биосистем [1, 2]. Итак, аналогом проявления (образования) голограмм в организме является специфическая динамика хромосомных и других знаковых клеточно-тканевых структур, которые можно рассматривать как амплитудно-фазовые рассеивающие квантованно-динамичные квазиголографические и семантические образования. Одним из законов построения их являются градиенты интенсивности результирующих электромагнитных спекл-полей. Известно, что спекл-поля характеризуют степень модуляции волновых когерентных полей. Для упрощения выкладок ограничимся случаем линейной передаточной функции (7)
Подставив выражение (7) в соотношение (6), получим математическую модель амплитудного пропускания клеточно-голографической структуры (8)
Проанализируем
каждый член уравнения (8). Первый член (обозначим
его Второй член
уравнения (8) описывает автокорреляцию послойно
меняющегося амплитудно-фазового фрагмента
“портрета” организма. Обозначим его через
Тогда, при таком обозначении, амплитудное пропускание квазиголограммы можно выразить в виде (9)
Проходя через такую систему квазиголографического фильтра-транспаранта, когерентная волна будет “считывать” имеющуюся на нем информацию как с обычной голограммы. Однако в отличие от обычного считывания, известного в технике голографии, считывание в живом и динамичном организме, вероятно, имеет квантованный по времени характер с возвратами (за счет возвратной памяти в спектрах ФПУ, см. выше), также как квантованными являются движения молекулярных и надмолекулярных структур организма. Последнее отчетливо прослеживается в нелинейной динамике ДНК, хромосом, рибосом и коллагена in vitro - in vivo, по крайней мере, в рамках явления ФПУ [1, 2]. На уровне органов биосистемы квантованность - это прерывистые движение глазного яблока. Квантованность движения субклеточных органелл проявляется в так называемом ядерном циклозе, при котором клеточные ядра нейронов или кариоплазма вращаются. У нейронов вращение ядра происходит по часовой стрелке, затем оно на некоторое время прерывается и продолжается уже в противоположном направлении [8]. В таком случае,
даже при непрерывном прохождении восстанавливающей
референтной волны всегда возможно квантованное по
времени когерентное считывание и когерентный
квантованный прием обогащенного информацией
волнового фронта, прошедшего через такую
биоквазиголограмму. В моменты кратковременных
остановок вращения при прохождении когерентной
волной (10)
Также как и на этапе регистрации ограничимся параболическим приближением: (11)
Восстанавливающая
когерентная волна (10), проходя через
новообразованный слой квазибиоголограммы,
дифрагирует на ее тонкой амплитудно-фазовой
структуре. Образующуюся после этого информационную
волну можно описать выражением, подобным выражению
(1), с той лишь разницей, что в фазовом члене
будет присутствовать расстояние (12)
причем, (13) где Подставив амплитудный коэффициент пропускания (9) в соотношение (12), получим четыре члена, из которых первые два определяют закон построения тонкой биоструктуры, а реконструированному изображению, как волновой схеме потенциальной биоструктуры, соответствуют два последних члена. Последние проанализируем отдельно. Вначале рассмотрим третий член, которому соответствует амплитудное пропускание (14)
Подставляя это выражение в соотношение для амплитуды дифрагированной волны, получим: (15)
Подставив в
данное уравнение для (16)
Это выражение описывает амплитуду дифрагированной волны, появление которой обусловлено той частью структуры биоголограммы, которая приводит к реконструкции основного (или истинного) волнового изображения того, каким должна быть “планируемая” биоструктура. Для получения ее точного пространственного прообраза с максимально резким изображением, биосистеме необходимо начинать строительство потенциальной структуры на расстоянии, при котором исчезнет квадратичный фазовый член в уравнении (16), что произойдет при соблюдении следующего условия
Отсюда следует, что расстояние основного изображения от биоголограммы определится по соотношению (17)
На расстоянии
представляет одну
из возможных форм записи так называемой функции
Дирака
Обозначая в
уравнении (16) выражение перед интегралом через
При Теоретический анализ биофизики управляющих генетико-волновых процессов, проводимый в настоящей работе, а также ранее [1, 2], приводит к предположению о том, что многоклеточные организмы обладают высокой избыточностью и помехозащищенностью волновой генетической информации по типу голограмм. Вероятно, хромосомно-водный континуум биосистемы является когерентным голографическим фракталом, где волновая информация многократно (избыточно) повторена в разных пространственных масштабах. Отсюда становится понятным, почему основная часть генома биосистем (гетерохроматин) устойчива к повреждающим воздействиям. Она голографична. Другая, малая часть генома (эухроматин), работает, вероятно, на других информационных принципах, а именно в рамках семантико-лингвистических построений последовательностей нуклеотидов ДНК и РНК [2]. И поэтому повреждения эухроматина, как правило, бывают летальны. Это и понятно. Например, замена только одной буквы в генетическом “тексте” может полностью поменять смысл “слова”, как это имеет место при повреждениях генов ферментов. То же относится и к расстановке “знаков препинания” в ДНК-“текстах”. Однако и здесь роль внутренних и внешних для биосистемы физических полей значительна. Одной из областей их работы являются смысловые ориентации выбора омонимов кодонов-антикодонов информационных и транспортных РНК, которые в свою очередь, определяются контекстными перестройками хромосомно-волнового континуума биосистем, а они, перестройки, в какой-то мере, зависят от волнового окружения земли, выступающего как мегаконтекст (фон) [2]. И в этом видится объединяющий фрактальный принцип в знаковой системе геном-поле. Если вернуться к нашей идее, что хромосомы высших биосистем могут работать также и на принципах квантовой телепортации знаковых фотонных образов, то можно сказать следующее. Геном нелокален трижды. Один раз по вещественному измерению, второй и третий - по волновому. Первое измерение нелокальности имеет вещественный характер. Это нелокальность голограмм и квазиголограмм хромосомного континуума. В этом же и фрактальность генома, поскольку любая голограмма - фрактал, т.е. информация в ней (записанный образ) самотождественна в любом ее фрагменте, не меньшем, чем длины волн записи. Второе измерение (волновое) - нелокальность волновых фронтов, считанных с таких голограмм. Каждая область фронта также фрактально повторяет образ, считанный с голографической решетки. И наконец, третье измерение (тоже волновое) - это квантовая нелокальность генома, позволяющая находиться в мгновенном электромагнитном информационном контакте миллиардам клеточных ядер, благодаря чему организм является суперкогерентной системой. Нам предсталяется, что прием и передача биоголографической (и радиоволновой) информации может происходить в квантованном по времени когерентно-импульсном режиме посредством квазизатворов. Если говорить о фотонных излучениях хромосом, то в качестве квазизатворов (или шире - переключателей с одной голограммы на другую) могут выступать холестерические послойные жидкокристаллические голограммы хромосомного континуума. Его холестерики посредством переориентации директоров меняют направление поляризации проходящего через них света. Такие регуляторные директора, расположенные ортогонально друг к другу и ортогонально к распространяющемуся свету, могут пропускать или не пропускать его за счет поворотов плоскостей поляризации, определяемых направлениями директоров. Когерентно-импульсный режим работы хромосомного континуума как лазерного излучателя может осуществляться по двум возможным путям. Первый основан на поворотах плоскостей поляризации света директорами хромосомных холестериков ДНК, а второй связан с известным явлением по захвату и накоплению фотонов в слоисто-фрактальных структурах, в том числе вероятно, и в жидких кристаллах ДНК. При такой ситуации может происходить лазерная накачка фотонов молекулами ДНК с их последующим квантово-пороговым импульсным отстрелом. Видимо, именно этот эффект мы наблюдали ранее, фиксируя необычные трапециевидные временные автокорреляционные функции при динамической лазерной спектроскопии светорассеяния ДНК и хромосом [1]. В качестве определенного подтверждения этих положений нами были проведены предварительные экспериментальные исследования по изготовлению и функционированию специальных ДНК-слоистых устройств типа “сэндвич” сложного состава, включающего кроме ДНК определенные биометаболиты. Такие устройства при взаимодействии с особым образом поляризованным лазерным светом со специальной юстировкой к вектору распространения луча лазера, оказались способными конвертировать фотоны в биоактивные радиоволны. Такие радиоволны меняют структуру воды, вводя в нее некую часть информации с ДНК. После этого вода оказывает четко выраженное действие на рост и развитие растений. Другой иллюстрацией биоактивности ДНК-промодулированных радиоволн является эффект оживления невсхожих семян растения Arabidipsis taliana, взятых из чернобыльской зоны и получивших высокую дозу гамма-облучения. Последний эксперимент служит некоторым свидетельством в пользу нашей гипотезы о квантовой нелокальности геномов высших биосистем, поскольку какая-то часть информации с ДНК передавалась параметрически связанной конверсией фотоны-радиоволны на поврежденный геном Arabidipsis taliana и регенерировала его. Это наблюдение также подтверждает полученные нами ранее данные о восстановлении хромосомного аппарата радиационно поврежденных семян пшеницы и ячменя с помощью радиоэлектронных устройств, моделирующих радиоволновые процессы в хромосомах [1]. Таким образом, мы предложили более развитую био-физико-математическая модель волнового управления ростом и развитием многоклеточных организмов, дополняющая и расширяющая первичные, выдвинутые нами ранее [1, 2], представления в этой области. ЛИТЕРАТУРА 2. Гаряев П.П. Волновой генетический код М.: Издатцентр. 1997. 3. Прангишвили И.В., Ануашвили А.Н., Маклаков В.В. Закономерности проявления подвижного объекта / Сб. трудов ИПУ РАН. М. Вып. 1. С.7-10. (1993). 4. Livolant F. Condensed forms of DNA in vivo and in vitro // Physica A. 1991.V.176. P.117-137. 5. Милер М. Голография. Л.: Машиностроение (1979). 6. Гаряев П.П., Тертышный Г.Г., Готовский Ю.В. Трансформация света в радиоволны. III международная конференция “Теоретические и клинические аспекты применения адаптивной резонансной и мультирезонансной терапии”. “ИМЕДИС”. Москва. 18-20 апреля 1997г. с.303-313. (1997). 7. Гаряев П.П., Тертышный Г.Г., Лощилов В.И., Щеглов В.А., Готовский Ю.В. Явление перехода света в радиоволны применительно к биосистемам. Сборник научных трудов “Актуальные проблемы создания биотехнических систем”. Выпуск 2. МГТУ им. Н.Э.Баумана. Академия Медико-Технических Наук РФ. Москва. С.31-42. (1997). 8. Движение немышечных клеток и их компонентов. Академия Наук СССР. Л.: Наука, 1977. С.105, 167. 9. Einstein A., Podolsky B., Rosen N., Can quantum-mechanical description of physical reality be considered complete? // Phys.Rev. 1935. V.47. P.777-780; Bouwmeester D., Pan Jian-Wei, Mattle K., Eibl M., Weinfurter H., Zeilinger A. Experimental quantum teleportation // Nature. 1997. V.390. P.575-579. |
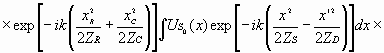
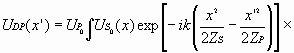
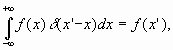 следовательно, (19)
следовательно, (19)